
専任教官の研究紹介
集積型錯体の構築とスピン状態制御 中島 覚
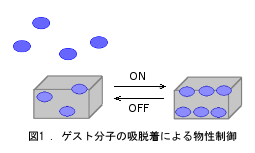
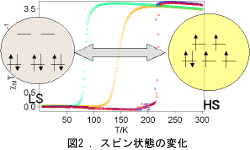
多孔性集積型錯体は種々の物性発現の源となる金属原子と分子設計性を有する有機配位子を持つとともに、 空孔が存在するため、これを利用した物性制御が可能となる(図1)。 一方、鉄二価錯体はd電子を6個持つが、配位子場の強さに依存して高スピン(HS)状態か低スピン(LS)状態を取る。 中間の配位子場であれば温度等によりスピン状態が変化する(図2)。 溶液中だとなだらかに変化するのみだが、結晶中では錯体間の相互作用に依存する。場合によってはヒステリシスを伴う。 従って、どのように錯体を集積するかが重要となる。
anti-gauche異性を持つ1,2-ビス(4-ピリジル)エタン(bpa)を架橋配位子として用いることにより、その異性を反映した多彩な集積構造 (1D直鎖構造、2Dグリッド構造、相互貫入構造)を得た。そのままでは高スピン状態のみを示したが、空孔に有機分子を導入することにより スピンクロスオーバーを実現した。そして、集積構造の違い、ゲスト分子の違い、アニオンの違いによりその挙動が制御できることが分かった。 さらに、この系においてメスバウアーパラメータと転移温度との間に密接な関係が見られた。
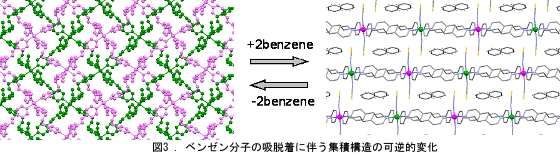
架橋配位子を1,3-ビス(4-ピリジル)プロパン(bpp)に変えることにより、2枚のグリッド構造が二次元的に相互貫入し、 それらが積み重なった大変珍しい集積構造を得た。合成溶媒を変えることにより、ベンゼンを包接した1D鎖状構造を得た。 この系では、ベンゼン分子の可逆的な吸脱着がみられるとともに、可逆的な集積構造の変化も見られた(図3)。 アニオンを選択することにより、集積構造の可逆的な変化に伴いスピン状態をスイッチさせることに成功した。
1) T. Morita, S. Nakashima, K. Yamada, and K. Inoue, Chem. Lett., 35, 1042-1043 (2006).
2) M. Atsuchi, H. Higashikawa, Y. Yoshida, S. Nakashima, and K. Inoue, Chem. Lett., 36, 1064-1065 (2007).
3) S. Nakashima, T. Morita, and K. Inoue, Hyperfine Interact., 188, 107-111 (2009).
4) M. Atsuchi, K. Inoue, and S. Nakashima, Inorg. Chim. Acta, in press (2011).
フルバレンで架橋した二核錯体の酸化的付加反応 中島 覚
環境放射能動態における微生物の影響に関する研究 稲田 晋宣
これまで、大学と公共下水道の接続部および角脇調節池の水の放射能を測定してきた。これまでの測定結果から、 公共下水道の放射能測定値は角脇調節池のものと比較して、全β測定値が高く、また周期的変動が確認されている。 環境放射能の変動は主に気象変化で議論されることが多いが、公共下水道は閉鎖系であり、気象変化の影響を受けにくい。 下水中の残渣の元素分析の結果、炭素元素(成分)を多く含んでいることが確認された。これは、有機物を豊富に含んでいることを示唆している。 また下水は保管中にpHが低下することが確認された(角脇水では確認されていない)。保管中は外部からの因子の混入はなく、 この現象は下水中に存在する微生物による影響であることが考えられ、この微生物の活動が下水の環境放射能の周期的変動にも何らかの 関与をしていると考え、微生物の影響について検討した。
環境水中の微生物について、固体培地上の生育を確認した。その結果、年間を通じて106~107 /mLのコロニー形成数を示し、 特に夏季に多く形成する傾向を示した。また水中の微生物種を確認するために、16s rDNAの部分配列を用いたDGGE解析を行った。 その結果、発現プロファイルにも変化が確認され、夏季には種類が増えていることが確認された。この結果、夏季には気温(ならびに水温)が 上昇し、水中の微生物の活動が活発となり活動する微生物の種類や数が増加していることが示唆された。
また下水中の微生物を定温培養すると、凝集する性質があることが明らかとなった。この凝集を行う時に水中の不純物を 巻き込んでいた。この現象は栄養が豊富な時には確認されず、栄養枯渇などの(特定の)条件により起こるものと思われる。 このような微生物活動の影響が環境放射能動態に何らかの影響を及ぼしている可能性がある。またこうした機能は環境中の金属元素などに 作用していることが考えられる。このような微生物の機能を解析することで、より有用な微生物の機能を発見できる可能性がある。 現在、環境中の微生物による影響やその機能について解析を行っている。
Proceedings of the Eleventh Workshop on Environmental Radioactivity, 136-140,
KEK Proceedings, 2010-8 (2010)
微生物における金属元素に対する影響 稲田 晋宣
植物に存在する還元酵素類の機能・構造の解明 松嶋 亮人
ある化合物は炭素を中心し、4種類の異なる官能基A,B,C,Dが結合しているとします。 その化合物を鏡に映すと全く同じものが映っているように見えますが、元の化合物と映った化合物は右手と左手が重なり合わないのと 同様に異なる性質を持つ化合物です。このような関係を鏡像異性体と呼びますが、これらを化学合成で作り出すことは非常に困難です。
一方、生物が持つ酵素は鏡像異性体を認識し、反応させることができます。この性質を利用して、植物由来の酵素を用いてケトンに共役した 炭素=炭素二重結合を還元することを研究の目的としています。
![]()
黒瀬川支流に形成されたバイオマットを構成する微生物の同定 松嶋 亮人
利用グループの研究紹介
プレゴン還元酵素の立体選択性は蛋白質複合体が決定する
理学研究科 数理分子生命理学専攻
泉 俊輔
我々の研究室では、本センターの松嶋先生と共同で、植物培養細胞を用いた物質変換について研究を行っている。 高等植物には3種のエノン類を還元する酵素が存在することが知られており、そのうちのPulegone還元酵素は(R)-Pulegone(1)を (1R,4R)-Isomenthone(2)と(1R,4S)-Menthone(3)とにジアステレオマー選択的に変換する。しかしながら Pulegone還元酵素を大腸菌を用いて 大量発現し、この酵素に(R)-Pulegone(1)を反応させたところ、 2と3の生成には立体選択性が見られなかった。 一方、大腸菌で発現させたPulegone還元酵素にタバコ培養細胞の蛋白質溶液を加え、(R)-Pulegone(1)を反応させたところ、 (1R,4R)-Isomenthone(2)が60%のジアステレオマー選択性(d.e.)で生成した。我々は、Pulegone還元酵素の立体選択性を決定する因子を タバコ培養細胞の蛋白質溶液中から同定し、その因子のPulegone還元酵素との結合位置と立体選択性発現のために、この因子がPulegone還元酵素に及ぼす影響を調べた。
まず、大腸菌で発現させたPulegone還元酵素をTresyl-Toyopearl担体に固定化し、タバコ培養細胞の蛋白質溶液のうち、 Pulegone還元酵素と結合した蛋白質を溶出した。その結果、Pulegone還元酵素にはβ-D-Glucosidase(32 kDa)が結合することがわかった。 次に、Pulegone還元酵素にβ-D-Glucosidaseを加えて(R)-Pulegone(1)の還元における立体選択性の変化を調べた。 その結果、β-D-Glucosidaseを加えることにより、d.e.が34 %まで上昇することがわかった。ここでβ-D-GlucosidaseとPulegone還元酵素を クロスリンクし、β-D-Glucosidase の結合位置を調べた。その結果、β-D-Glucosidase はPulegone還元酵素の活性部位付近のループ構造 (Gly255~Asn270)に結合することがわかった。
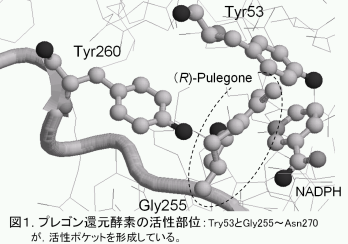
以上の結果より、Pulegone還元酵素は、図1に示すように、Pulegone の活性部位へ異なる方向から組み込まれることによって、 (1R,4R)-Isomenthone(2)および(1R,4S)- Menthone(3)の生成の立体選択性が生じることが明らかになった。 すなわち、Pulegone還元酵素に結合蛋白質が結合すると、活性部位付近のループ構造を押し上げて、(1R,4S)-Menthone(3)生成に有利な基質導入 が困難となることが示唆された。
高等植物における生体防御機構-アポトーシス実行の初期過程
理学研究科生物化学研究グループ
平田敏文・泉 俊輔・芦田嘉之
生き物は外界からの刺激に対して自らの身を守るために生体防御反応がプログラムされている。そのプログラムの中には自らの細胞の一部を死滅させ、それによって自らを防御する防御機構(=アポトーシス)があることが知られている。我々は植物細胞に化学的ストレスを負荷すると、その細胞がアポトーシス様の細胞死に至ることを見出し、アポトーシス発現の分子機構の解明を計っている。ここでは、アポトーシス誘導の初期過程について、我々の最近の研究結果を紹介する。

化学的ストレスとしてゲラニオールを投与したタバコ培養細胞の細胞質画分を粗酵素液として、配列既知のタンパク質を加水分解させた。得られた消化断片を分析すると、すべての消化断片はグルタミン酸をC末端とすることが明らかになった。そこで、グルタミン酸p-ニトロアニリドの加水分解活性を指標としてプロテアーゼの精製を計り、SDS-PAGEで42 kDaと24 kDaにスポットを示す蛋白質を得た。次に、42kDaと24 kDaのバンドをそれぞれ切り出し、トリプシンでゲル内消化の後、ペプチド・マスフィンガープリント法により検索した。その結果、どちらのスポットから得られたペプチド配列も、発芽時に活性化されるタバコのシステインプロテアーゼの配列に一致した。しかし図1に示すように、24 kDaのタンパク質のトリプシン消化断片では、N末端から120残基までに由来する消化断片が現れなかった。また、24 kDaのプロテアーゼをMALDI TOF-MSにより測定したところ、その分子量は24200であった。これらのことより、このプロテアーゼは細胞質内で118番目のグルタミン酸までが失われて、活性型のプロテアーゼとなることが示唆された。このプロテアーゼを用いて、アポトーシスの初期段階で分解されるカルレティキュリンの加水分解を行った。その結果、カルレティキュリンはプロテアーゼ投与後10分で、C末端から3520Daのフラグメントを分解産物として与えた。このことより、細胞内でカルレティキュリンはゲラ二オール投与時に活性化されるシステインプロテアーゼによって、C末端から29残基が『削り込まれる』ことが示唆された。これらの結果より、アポトーシスの誘導過程において、植物細胞内ではシステインプロテアーゼが活性化され、これによってカルレティキュリンのC末端の小胞残留シグナルを『削りこむ』ことが明らかになった。カルレティキュリンは核内移行シグナルを持っているので、小胞残留シグナルを削りこまれたカルレティキュリンは核内移行し、ヒストンと結合することにより、ヌクレオソーム単位でのクロマチンの断片化に寄与しているものと考えられる。植物におけるアポトーシスの分子機構の完全解明のためには、さらなる研究が必要である。
福島第一原発事故由来放射性セシウムのスギ樹葉中の分布
サステナブル・ディベロップメント実践研究センター 田中 万也
新しいがん治療法開発への応用を目指した染色体末端テロメアの機能解析
先端物質科学研究科 分子生命機能科学専攻 上野 勝
1 次繊毛における中枢性摂食受容体MCHR1 の機能解析
総合科学研究科 生命科学領域 斎藤 祐見子


